Physiologie Végétale :
Partie 1 : nutrition et métabolisme.
Chapitre 3 : La nutrition azotée.

Télécharger le document (toute la partie 1) sur le serveur FTP ou sur le serveur web (HTTP).
I Les différentes formes de l’azote présent dans le minéral.
L’azote sous forme organique ou minérale représente 1 à 5% de la matière sèche. On trouve l’azote dans les protéines qui sont caractérisées par un taux moyen de 16% de cet élément. Le dosage de l’azote permet un dosage des protéines. Le pourcentage de protéines est : %N*6,25 (6,25 représente le coefficient de Kjeldahl : 100/16=6,25). C’est la méthode la plus précise mais pas la plus utilisée. On trouve l’azote dans les acides nucléiques, les coenzymes, les vitamines, les hormones… Quand l’azote est sous forme minérale, c’est sous une forme ionique comme NH4+ ou NO3–.
II Les différentes formes d’azote disponible dans la biosphère.
A L’azote atmosphérique.
Il représente 78% de l’air, c’est donc la principale source. Toutefois, seules quelques plantes qui vivent en symbiose (bactéries ou algues) sont capables d’utiliser directement l’azote atmosphérique.
B L’azote du sol.
L’azote a cinq électrons sur sa couche externe dont trois sont célibataires et peuvent donc établir des liaisons covalentes. Le nombre d’oxydation de l’azote varie de –3 à +5.
On trouve l’azote minéral sous trois formes : NO3–, NO2–, NH4+. L’azote organique se trouve dans des molécules complexes qui peuvent être des protéines ou des acides aminés. Ces molécules se trouvent surtout au niveau de l’humus. La matière organique décomposée va être à l’origine de l’azote utilisé par la plante.
1 Transformation rapide, décomposition, humidification, minéralisation.
On trouve différents types de décomposition :

La transformation rapide nécessite un processus microbien puis une phase physico-chimique (cette phase entraînera la libération de substances nutritives). Certains composés (10%) sont difficilement dégradables et passent donc par une phase de dégradation lente.
2 La transformation lente.
Elle a surtout lieu pour les composés comme la cellulose, la lignine. Ces composés sont lentement oxydés, polymérisés (processus chimique). Il y a alternance de dessiccation et d’humidification et intervention de micro-organismes (grosses molécules → très grosses molécules → minéraux). L’humus est une fraction colloïdale de la matière organique obtenue par synthèse microbienne (physico-chimique) à partir de matière organique végétale.
3 La minéralisation.
La protéolyse : les protéines sont dégradées en acides aminés ou en amides.
Ammonisation : l’urée va donner de l’ammoniac, des sels ammoniacaux, des carbonates d’ammonium (CO3(NH4)2). Les sels d’ammonium vont subir la nitrification.
La nitrification transforme l’ammoniac et le NH4+ en ions nitrate par 2 oxydations successives. NH4+ (-III) → NO3– (+V). La somme des nombres d’oxydation donne la charge de l’ion.
La nitrosation : l’acide nitreux et le nitrite. 2 NH4+ + 3 O2 → 2 NO2– +2 H2O + 4H+ (ΔG’°=-542kJ/mol). Il y a production d’énergie. La réaction et réalisé par les bactéries au niveau du sol. Ces bactéries sont du genre Nitrosomonas. Les ions NO2– sont des ions toxiques.
La nitratation : l’acide nitrique, le nitrate. 2 NO2– + O2 → 2 NO3– (ΔG’°=-155kJ/mol). Cette réaction est réalisée par les bactéries du genre Nitrobacter qui sont des chimiolithotrophes. Ces bactéries sont autotrophes pour le carbone et utilisent l’énergie pour assimiler ce carbone.
III Assimilation de l’azote minéral par la plante.

Quel que soit l’ion de départ, il passe toujours par le stade NH4+. Bien que les végétaux puissent utiliser l’ammoniaque, ils se servent surtout de l’ion NO3– : c’est la réduction des nitrates. L’assimilation de l’azote minéral est réalisée dans les racines, dans les bourgeons ou dans les feuilles.
Dans les racines, NO3– donne des amides et des uréides transformés puis transportés par le xylème.
Dans les tiges et les feuilles, NO3– donne des amides qui se sont transportés par le phloëme. Les enzymes sont : la nitrate réductase (NO3– → NO2– 2e–) ou la nitrite réductase (NO2– → NH4+ 6e–).
A Assimilation des nitrates.
1 La nitrate réductase. NO3– → NO2– (2e–).
Cette enzyme est une flavo-métallo-protéine cytoplasmique. E’°= +0,46. Dans la racine, il y a utilisation de NADPH ou de NADH. Cette réaction inhibée par le NH4+ et a des effets inducteurs par le NO3–. L’enzyme est sensible à la lumière et aux hormones (cytokinines).
b La nitrite réductase. NO2– → NH4+ (6e–).
L’enzyme est une métallo-protéine qui renferme du fer dont le rôle est de catalyser une série de réactions dont l’intermédiaire est mal connu. Dans le stroma, il y oxydoréduction avec le donneur (ferrédoxine ou NADPH). Les plantes préfèrent absorber l’ion NO3–.
BAssimilation de l’azote.
On a deux possibilités majeures :
1 La glutamate déshydrogénase (GDH).
Il existe du GDH mitochondrial et chloroplastique. Ce produit se transforme :
α-cétoglutamate + NH4+ ↔ Glutamate
COOH-C=O-CH2-CH2-COOH //// COOH-CNH2-CH2-CH2-COOH
C’est le phénomène d’amination réductrice. L’enzyme fonctionne avec le NADPH ou le NADH selon sa localisation.
2 La glutamine synthase (GS-GOGAT).
GS : glutamine synthétase ; GOGAT : glutamate synthase.
glutamate + NH4+ → glutamine
glutamine + α-cétoglutarate → glutamate + glutamate.
Le Glutamate va vers le métabolisme.
COOH-CNH2-CH2-CH2-C(=O)-NH2 : formule de la glutamine.
La première en réaction demande l’utilisation d’ATP. Il est sous sa forme cyclique, dans le chloroplaste et le cytoplasme de tous les végétaux supérieurs. Dans les nodules (dans le cas des symbiontes) et chez les cyanobactéries, on a, comme pour le GDH, des iso-enzymes.
La réaction de trans-amination (à partir de la glutamine et du glutamate) donnera toujours des amides, qui, celle-ci, cède un ammonium à des acides cétoniques. On a alors différentes familles d’acides aminés :
- glutamate : 5C
- aspartate : 4C (il vient de l’AOA). AOA → CCO-CNH2-CH2-COOH
- Pyruvate : 3C. Pyruvate → Alanine (COOH-C(NH2)-CH3).
- Sérine : CHOH à la place de l’alanine.
- Shikimate :
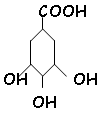 Cette forme est à l’origine de tous les AA aromatiques.
Cette forme est à l’origine de tous les AA aromatiques.
IV Utilisation de l’azote atmosphérique.
Les plantes utilisant cet azote sont dites diazotrophes. Elles possèdent une enzyme particulière (la nitrogénase). C’est une enzyme de micro-organismes.
A Les différents types de micro-organismes fixateurs d’azote.
les fixateurs libres.
Ils vivent dans le sol et quand ils meurent, ils donnent l’humus. Cet humus sera utilisé par la plante.
- Les hétérotrophes au carbone : On trouve les bactéries aérobies comme Azobacter. On trouve aussi des bactéries anaérobies de type Clostridium.
- Les autotrophes au carbone : on trouve des photo-bactéries anaérobies a un photosystème comme celles de type Rhodospirillum. On peut aussi trouver des cyanobactéries comme Nostoc et Anaboena.
Azospiryllum est un fixateur libre de l’azote qui vit dans la rhizosphère des pieds de maïs.
Les fixateurs symbiotiques.
On trouve les hétérotrophes au carbone comme, les rhizobiacées (anaérobies), par exemple, les légumineuses. On pouvait aussi, trouver des actinomycètes (Frankia, qui parasite les plantes ligneuses). Parmi les autres opprobres au carbone, on trouve au des cyanobactéries comme Nostoc, Anaboena mais aussi des hépatiques, des fougères, cycadacées, gunnera…
Définition d’un nodule : c’est un organe différencié dans la plante, où se développent les bactéries fixatrices d’azote. Les bactéries se transforment en bactéroïdes (qui fixent l’azote) de façon irréversible.
B/ La nitrogénase (ATP dépendante).
Cette enzyme est composée de six sous-unités. Ces six parties forment une protéine ferrique à deux sous-unités (dinitrogénase réductase azoferredoxine). C’est elle qui utilise l’ATP et qui transfert les électrons à l’autre sous-unité : la dinitrogénase (protéine qui contient du fer-molybdène). Elle qui fixe l’azote en formant l’ion NH4+. Elle reçoit les électrons de la première partie protéique et fixe l’azote. Le résultat de cette activité représente 10% des protéines solubles dans les bactéroïdes.
La réduction de l’azote atmosphérique.
Le pouvoir réducteur vient de la ferrédoxine ou d’autres réactions comme la décarboxylation oxydative du pyruvate, cette réaction demande de l’ATP et produit de l’hydrogène. L’ATP vient de la photosynthèse ou de la phosphorylation oxydative. L’azote est transformé en matières organiques comment précédemment.
Le problème de la régulation : la dinitrogénase est très sensible à l’oxygène et est inhibée irréversiblement par des traces d’oxygène. Dans le cas des organismes anaérobies, il n’y a pas de problème.
Chez les algues bleues, la compartimentation de la nitrogénase, dans les hétérocystes empêche ce problème (photosystèmes).
Dans le cas des fixateurs symbiotiques, libres et aérobies, l’oxygène est piégé par une respiration amplifiée. Dans le cas particulier des symbiotiques, on a en plus des métallo-protéines (chromoprotéines) qui contiennent un hème : la leghémoglobine qui piège l’oxygène. Cette protéine représente 40% des protéines des nodules de légumineuses. La leghémoglobine fait partie des nodulines, cette protéine est synthétisée par la plante seulement dans les nodules. L’azote ammoniacal et les nitrates sont des répresseurs pour la plante. Dans la nature on n’est jamais en conditions d’inhibition.
Pourquoi la GDH ne peut elle pas remplacer le cycle GS/GoGat ds le processus général de la nutrition azotée?
Très bien détaillée le cours