Microbiologie :
Chapitre 1 : Structure des micro-organismes.

Télécharger le document sur le serveur FTP ou sur le serveur web (HTTP).
I Les bactéries.
A Généralités.
Les micro-organismes sont les plus petites unités biologiques fonctionnelles. Leur taille est comprise entre 0,01 et 10 µm.
Les différentes formes existantes :
- Les coques : ils sont sphériques. Les différents assemblages de coques proviennent de la division cellulaire.
- Les bâtonnets (bacilles) : ils sont droits (coli bacille), incurvés comme les vibrions (exemple : Vibrio cholerae). On peut aussi trouver des formes irrégulières et enflées aux extrémités (exemple : mycobacter, corybacter). Une autre forme possible est la forme coccobacille (exemple : Serratia marcescens).
- Les hélicoïdaux : on trouve les spirochètes qui pourraient être à l’origine des mouvements des premières cellules eucaryotes
- Les formes mycelliennes : ce sont des formes filamenteuses, comme les Actinomycètes. Les myxobacters ont des organes de fructification qui sont des ensembles de bactéries de forme similaires.
B Structure.
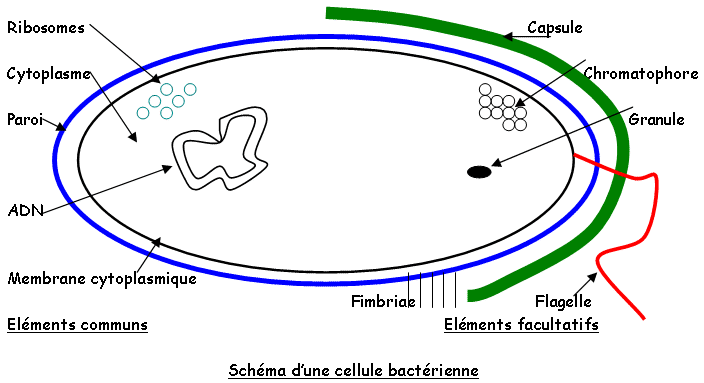
Les micro-organismes sont étudiés par fractionnement, par immunocytologie, ou au microscope électronique à balayage (MEB). On a un schéma type de bactéries avec des composants types comme les ribosomes, le cytoplasme, la paroi, la membrane cytoplasmique et l’ADN. On peut aussi trouver des composants facultatifs comme les granules, les réserves, les chromatophores, des fimbriae, des flagelles et des capsules.
1 La membrane plasmique.
Cette membrane a une structure classique, en double feuillet, mais moins rigide que les membranes eucaryotes car elles ne possèdent pas de stérols (sauf les mycoplasmes).
a Composition.
Cette membrane est composée de 30 à 40% de lipides (dont les glycérophospholipides) et de 60 à 70% de protéines.
b Rôle et fonction.
C’est une barrière semi-perméable qui permet le transport passif de certaines molécules grâce à des protéines canales.
Les protéines membranaires sont des enzymes permettant des biosynthèses de lipides bactériens ou de peptidoglycanes.
La membrane peut avoir un rôle respiratoire grâce à la présence de cytochromes dans cette membrane (un peu comme une mitochondrie).
2 Le cytoplasme.
Il contient 80% d’eau avec un pH compris entre 7 et 7,2 où sont dissous des sucres, des ions, des acides aminés… On y trouve aussi des inclusions de granules de polyphosphates ou de polyhydroxybutirate, des ribosomes associés à l’ARNm sous forme de polysomes (1000/cellule).
3 Le nucléoïde.
C’est le génome bactérien (ou chromosome bactérien). Il peut exister sous plusieurs copies en même temps. On assiste au phénomène d’amitose : il n’y a pas de synchronisation entre la division de l’ADN et celle de la cellule.
a Organisation.
Ce nucléoïde n’est pas isolé du cytoplasme par une membrane. Il est constitué par un double brin circulaire (refermé) d’ADN : c’est un assemblage en double hélice de deux chaînes antiparallèles et complémentaires de nucléotides. (A et G sont puriques alors que C et T sont pyrimidiques).

L’ADN bactérien est sans histones et est super-enroulé. Très souvent, ce dernier est ancré en un ou plusieurs points de la membrane plasmique. Par exemple, chez Escherichia coli, on trouve 5 millions de paires de base où 4300 gènes sont identifiés (60% du total). Cet ADN mesure 1mm quand il est déroulé et représente 10% du volume cellulaire.
b Rôle.
C’est le support de l’hérédité. Ce nucléoïde permet de lire le génome, de le transcrire en ADN (afin de synthétiser des protéines). Il subit aussi la réplication pour assurer la descendance.
c Biosynthèse de nucléoïdes.
La réplication est la synthèse d’un nouveau génome : c’est un mécanisme semi-conservateur et bidirectionnel.
Il apparaît une fourche de réplication grâce à une DNA-polymérase. Il y a phosphodiestérification entre l’amorce d’ADN et ce qui est lu dans le sens 3’→5’.
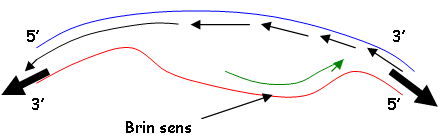
On parle de brin sens quand il y a réplication dans le même sens que celui de l’ouverture de la fourche de réplication.
Les fragments synthétisés sur le brin anti-sens sont reliés par une ADN-ligase.
Les deux brins sont synthétisés en même temps dans les deux sens.
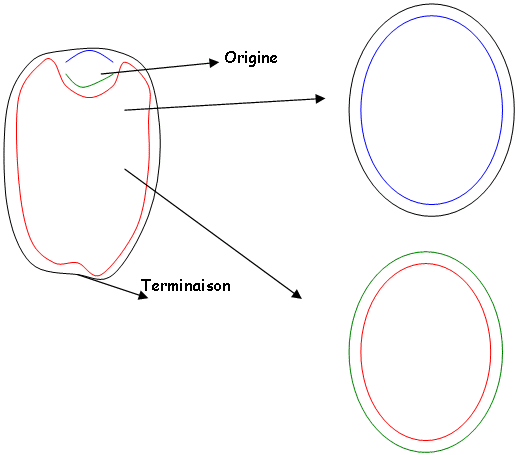
L’ADN-gyrase permet de désenrouler au point d’origine et suit le mouvement des fourches. Cette enzyme est inhibée la norobiocine (antibiotique).
d Les plasmides.
Définition : ce sont des molécules d’ADN double brin, circulaire, qui, extrachromosomique, ne constituent pas le génome bactérien. Ils ont une réplication autonome (un pouvoir infectieux), une petite taille et codent pour une information génétique non-indispensable. Ils peuvent infecter des bactéries ou être échangés entre elles.
Ces plasmides ont été découverts en 1952 sur Shigella dysenteriae.
Shigella résistante + E. coli saine → Shigella résistante
→ E. coli résistante
→ E. coli saine
Réplication et transfert : elle se fait de façon autonome selon le même processus que celui du nucléoïde. Ils ont la même vitesse de réplication : on peut donc avoir plusieurs plasmides en même temps dans une même cellule. Le transfert des plasmides se fait par conjugaison :
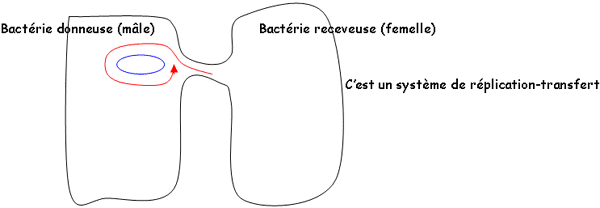
Propriétés : les plasmides sont des unités codantes. Ils donnent à la bactérie :
- la possibilité de synthèses spéciales
- une résistance à des antibiotiques : comme des enzymes qui dégradent les antibiotiques (par exemple : la ß-lactamase qui résiste aux pénicillines.
- Une pathogénicité. Chez E. coli, il y a synthèse d’entérotoxines qui provoquent des maladies. E. coli peut aussi gagner l’aptitude à se fixer sur une membrane.
- Un pouvoir infectieux
- Le plasmide F donne la possibilité de recombinaison génétique : il code pour la synthèse de pili sexuels pendant la conjugaison.
e La paroi.
C’est la plus externe, c’est elle définit la cellule procaryotique. De plus, celle-ci sert à la classification des micro-organismes. Elle représente 20% de la masse sèche. Elle a un rôle majeur dans la résistance à la pression osmotique et aux déformations. Cette coloration (ou non-coloration) est révélatrice d’une différence structurale de la paroi.
Organisation :
La coloration de Gram (en 1844). C’est une étape préliminaire pour la reconnaissance d’une bactérie. Cette coloration est réalisée en présence d’iode, puis on lave à l’alcool : on trouve alors deux cas distincts :
- les bactéries sont décolorées : ce sont des gram-
- les bactéries gardent la coloration : ce sont les gram+.
La paroi des gram+ est 1000 fois plus grande que celle des gram-. La paroi des grm+ est généralement composée de peptidoglycane et d’acide teïchoïque. Ce dernier représente 50% du poids de la paroi structurée. La paroi des gram- est composée, sur la membrane externe, de lipoprotéines, de LipoPolySaccharides et de trimères : c’est un réseau lâche.
Remarque : Les mycoplasmes ne sont ni l’un ni l’autre : ils n’ont pas de paroi. Les archéons ont une structure un peu différente.
Composition globale : Dans les parois de gram-, on trouve plus d’acides aminés et de lipides.
Les macromolécules spécifiques
L’acide teïchoïque : il est composé, soit de ribitol (CH2OH-(CHOH)3-CH2OH), soit de glycérol. Ils sont substitués avec du glucose et/ou de l’alanine. Cet acide peut avoir un rôle de reconnaissance antigénique (sérotypie). Ces acides sont ancrés, soit dans la membrane, soit dans le peptidoglycane (où ils sont pariétaux). Ils servent aussi à relier la membrane et la paroi. Plus le réseau formé est dense, plus la paroi est rigide.
Le LPS : chez les gram-, il constitue les feuillets externes de la paroi externe de cette bactérie. Ce LPS est composé en trois parties
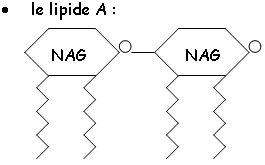
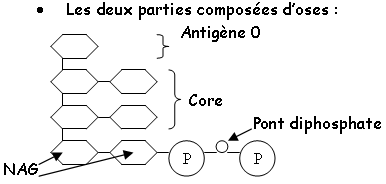
Le LPS a le même rôle antigénique que l’acide teïchoïque d’ancrage et de structuration de la membrane externe.
Les protéines de la membrane externe des Gram-.
On trouve deux grands types : les lipoprotéines qui sont des polypeptides qui lient la membrane externe au peptidoglycane et les protéines matricielles qui traversent la membrane externe et qui peuvent avoir un rôle dans le transport (porines) ou dans la réception des phages. Le peptidoglycane : (mureïne, glycocalix). Il a un haut poids moléculaire et est spécifique aux eubactéries. Il assure un rôle structurant (résistance et pression). Il donne donc la forme cellulaire et empêche la lyse par un milieu hypotonique.
Les constituants :
• Les osamines : ils sont composés de N-AcétylGlucosAmine ou d’Acide N-AcétylMuramique : c’est le composant de la paroi bactérienne. UDP-NAG + PEP + NADPH → UDP-NAM + NADP. Cette réaction est inhibée par la phosphonomycine. Chez les archéobactéries, il n’y a pas de muréine mais de la pseudo-muréine (pas de NAM mais du N-AcétylOsaminUronique).
• Les acides aminés :
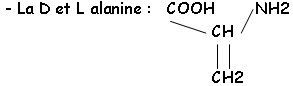
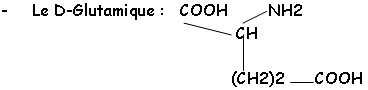

Cette chaîne est réticulée grâce au NAM et à son substitue en 3’ : il y a relation avec la L-Ala, la L-Lys ou le DAP, le D-Glu et la D-Ala.
Chez Staphylococcus aureus, on a L-Lys – (Gly)5 – D-Ala : c’est un maillage lâche qui lui donne sa forme sphérique.


Remarque : les mycoplasmes sont des parasites intracellulaires, donc dans un milieu isotonique. L’enveloppe comprend la paroi et la membrane.
Le rôle de la paroi : c’est une barrière (active chez les G- pour les transports). Le peptidoglycane assure la forme cellulaire. La couche la plus externe a des propriétés :
- antigéniques : des particules induisent la production d’anticorps : on obtient donc une définition de sérotypie bactérienne (grâce à l’acide teïchoïque chez g+, au LPS chez les Gram-.
- de fixation des phages (bactériophages = virus à bactéries). La recombinaison génétique à lieu grâce aux bactériophages. N’importe lequel de ces phages ne peut pas infecter n’importe quelle bactérie. Chez E. coli, le T4 se fixe sur le LPS, le λ se fixe sur la protéine qui transporte le maltose. Chez Bacillus suptilis, le f29 se fixe sur l’acide teïchoïque.
f Flagelles et pili.
Les flagelles.
Ils sont facultatifs, de nature protéique (la flagelline), sont des unités sphériques en hélice et sont ancrés dans la membrane plasmique. Ils per(mettent les déplacements microbiens : ils ont donc un rôle de chimiotaxie. Ils ont aussi des propriétés antigéniques : ils permettent de fixer les phages de type PBS1 sur Bacillus suptilis. On trouve différent type d’insertion des flagelles :

Le type d’insertion des flagelles peut être utilisé pour des reconnaissances bactériennes.
Pili (ou fimbriae)
Ce sont des éléments facultatifs de la cellule Gram-, ils sont de nature protéique comme les flagelles, mais en général plus courts. Ils procurent à la bactérie qui les portent, une capacité d’adhérence (souvent associée à la virulence de la bactérie). Ils ont un rôle dans la reconnaissance entre cellules donneuses et receveuses pendant la conjugaison. Ils sont aussi le site de fixation des phages comme le M13.
g La capsule.
C’est l’enveloppe supérieure la plus externe, souvent polysaccharidique (composée souvent d’acide hyaluronique). Cette capsule est associée à une virulence comme chez Klebsiella pneumoniae. Cette enveloppe entraîne un phénomène d’adhérence, mais elle masque aussi les sites antigéniques, et augmente la taille apparente de la bactérie pour résister à la phagocytose. Elle permet aussi la résistance à des conditions externes défavorables.
h Les endospores.
La sporulation est un phénomène induit par une carence nutritive. La germination nécessite une activation (choc thermique, forte variation de pH). La sporulation est une mise en l’abri du génome (le record de longévité est de 7500 ans) face aux conditions défavorables du milieu (dessiccation : perte d’eau du milieu).
Parfois, la formation des spores est associée à une libération de toxines. Par exemple, Bacillus thuregensis sécrète un cristal particulièrement herbicide.
La structure des spores :

C Classification.
La classification est réalisée par des
- Critères morphologiques (structuraux)
- Critères métaboliques (type respiratoire, condition de vie)
- Pili, endospores
C’est une approche phénotypique.
On peut aussi avoir une approche moléculaire pour se baser sur le génotype.
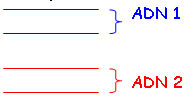
On va chauffer l’ADN pour le dénaturer. Ensuite, on observe si les ADN se recombinent ou non :
![]()
On a aussi une approche en pourcentage de G-C : ce pourcentage donne la température de dénaturation.
On peut aussi étudier les séquences d’ADN robosomiaux. Ils ont les propriétés d’être indispensables à la vie et de subir des variations pendant l’évolution. Ils permettent d’obtenir une distance génotypique. Exemple :
[table id=7 /]
TRES BON COURS
MERCI BINE POUR TOUT
merci
Très beau cours que je pense m’être très utile pour bientôt … Merci
J’aime bien ce cours ça le fait plaisir
J’aime bien ce cours c’est très intéressant