Biologie du Développement :
Chapitre 5 :
Le développement de la drosophile.

Télécharger le document sur le serveur FTP ou sur le serveur web (HTTP).
I La Drosophile melanogaster (la mouche du vinaigre).
Elle a été rendue célèbre avec l’avènement de la génétique classique au début du siècle et depuis 1980 par la biologie moléculaire. Grâce à cette mouche, on peut étudier le déterminisme moléculaire du développement.
La drosophile est une espèce qui présente de très nombreuses mutations spontanées qui affecte les caractéristiques morphologiques et physiologiques.
On peut provoquer des mutations par des agents chimiques ou ionisants. Les mutations peuvent affecter les mécanismes de l’orogenèse, du développement embryonnaire (larvaire ou pendant la métamorphose). Certains gènes séquencés chez la drosophile ont des équivalents dans les vertébrés supérieurs. Ils ont permis d’aborder l’étude du déterminisme moléculaire du développement chez les vertébrés supérieurs (entre autre, l’homme).
Le développement de la drosophile est très rapide : en 9 jours, on obtient un adulte. On voit donc rapidement de la mutation.
II Le cycle de vie de la drosophile.
A L’œuf centroleucyte (le vitellus est au centre de l’œuf).
La segmentation est dite superficielle. En effet, des noyaux se divisent au centre puis migrent vers l’extérieur et vont y former le blastomère. A 22°C, on a un développement très rapide.
Le développement embryonnaire dure 24 heures. La segmentation gastru-neuralation donne une larve de premier stade qui mue au bout de 24 heures et donne alors une larve de second stade qui va muer une nouvelle fois pour donner une larve de troisième stade. Pendant les trois stades larvaires, les organes mis en place pendant le développement embryonnaire vont croître. On assiste cependant à l’apparition d’une série d’ébauches indifférenciées : les disques imaginaux (è ils formeront souvent de l’adulte).
Après 48 heures, la larve commence à se métamorphoser. Elle conserve son tégument externe qui constitue le puparium (épaissi). C’est le stade de la pupaison. La dernière mue donne naissance à la pupe immobile ou l’on reconnaît les contours du corps et les appendices de la future mouche.
Le processus de la métamorphose est induit par des hormones stéroïdes (ce sont toutes des hormones de mue). Ce processus dure quatre jours. La larve sera en parti détruite par histolyse. Les images seront reconstituées par les disques imaginaux.
A l’issu de ces quatre jours, on se débarrasse du puparium. On a ensuite la ponte.
B L’orogenèse.
Les ovaires sont constitués de plusieurs tubes ou ovarioles qui contiennent les chambres contenant les œufs (chambres ovariennes). Ces chambres sont composées de seize cellules issues de la lignée germinale (cellules polaires).
On a quinze cellules nourricières et un oocyste. La chambre ovarienne est entourée par une couche de cellules folliculaires. Des ponts cytoplasmiques provenant d’une cytodiérèse incomplète permettent l’accumulation, dans l’oocyste, des métabolites nécessaires aux premiers stades de développement Des ponts cytoplasmiques provenant d’une cytodiérèse incomplète permettent l’accumulation, dans l’oocyste, des métabolites nécessaires aux premiers stades de développement, mais également à la répartition d’ARNm et de protéines qui sont des déterminants maternels.
Les cellules folliculaires produisent des composants du cytoplasme qui déterminent l’axe dorso-ventral. Les deux axes sont donc définis pendant l’orogenèse par des déterminants maternels.
Autour de l’orogenèse, les cellules sont agencées en palissade et forment ainsi la membrane vitelline et un chorion cireux et imperméable.
Les cellules nourricières et folliculaires sont responsables de la mise en place des axes embryonnaires par répartition polarisée des déterminants moléculaires maternels.
C Le développement embryonnaire.
La segmentation : les cycles cellulaires sont très brefs.
Après la fécondation qui est intérieure (dans la spermathèque), le développement commence par une série de huit divisions nucléaires synchrones qui se succèdent de huit à dix minutes sans formation de cellules individualisées. Ces noyaux constituent des énergides. Les 256 noyaux (ou énergides) formés vont migrer vers la périphérie de l’ooplaste (cytoplasme de l’oocyte) pour former le blastoderme syncytial (tissu plurinucléé).
A la neuvième division de la segmentation, des plasmes polaires vont s’individualiser en région postérieure (région de cytoplasme). Cinq ou 6 noyaux vont migrer puis pénétrer dans les plasmes et se viser pour donner une trentaine de cellules : les cellules polaires qui seront à l’origine du germanium. Après les quatre divisions synchrones syncytiales, on a treize divisions. Celles-ci permettent la formation des membranes cellulaires qui se forment de la périphérie vers le centre : c’est la cellularisation complète (d’une durée d’à peu près deux heures et demi). Le blastoderme est dit cellularisé.
Durant la transition entre 2 blastodermes, les gènes zygotiques commencent à s’exprimer, prenant le relais des déterminants maternels.
D La gastrulation.
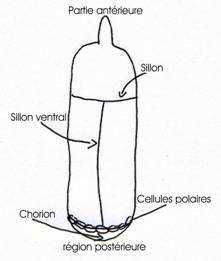
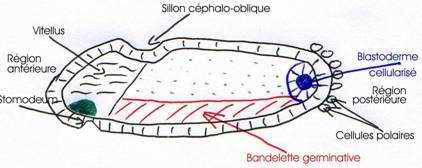
Le long de la ligne médiane ventrale, une bande de cellules s’invagine et forme le sillon ventral, puis un cordon ventral qui donnera le mésoderme accolé au futur ectoderme.
Ces deux formations constituent la bandelette germinatique, mise en place en vingt minutes.
Cette bandelette s’étend sur les deux tiers de la longueur de l’embryon et a pour limite antérieure, le sillon céphalique oblique qui se forme de façon superficielle et transitoire au même moment, donc, dès le début de la gastrulation.
L’ensemble ectoderme plus mésoderme forme la bandelette germinative.
On a ensuite deux épaississements du blastoderme qui constitue les ébauches endodermiques de l’intestin moyen antérieur et postérieur qui seront repoussés à l’intérieur de la formation par deux évaginations d’origine ectodermique.
On a une évagination à l’avant du système céphalique : le stomodeum qui donnera la bouche (œsophage) et le proctodeum qui donnera l’anus et l’intestin postérieur.
Les mouvements morphogénétiques de la gastrulation vont amener la bandelette germinative à s’enrouler autour de l’extrémité postérieure par un mouvement postérodorsal : une extension de cette bandelette qui est optimale en six heures.
Quand l’extrémité atteint le sillon céphalo-oblique, ce mouvement entraîne l’invagination du proctodeum qui se développe à l’intérieur des cellules polaires et du geme.
Entre 4 et 7 heures, de grosses cellules migrent, à partir de l’ectoderme, de part et d’autre du sillon médian : ce sont les neuroblastes.
Ces neuroblastes vont donner naissance à deux cordons nerveux entre l’ectoderme et l’endoderme selon un patron répétitif qui rappel les métamères de l’embryon.
Les sillons segmentés caractérisants la métamérisation apparaissent vers la 7ème heure sur la face ventrale, au moment ou le sillon céphalique disparaît.
On distingue 3 segments pour la tête, 3 pour le thorax et 8 pour l’abdomen. Entre 9 et 10 heures, on assiste à un retour en arrière de la bandelette germinative (rétraction de cette bandelette) qui place l’orifice du proctodeum à sa position définitive (anus) et les segments postérieurs se trouvent alors dans l’alignement axial du corps.
Le système nerveux subit ce phénomène de rétraction et lorsqu’il est dans sa position ventrale, une mince couche de cellules amnio-séreuses va recouvrir le vitellus exposé dorsalement après la rétraction de la bandelette germinative. On a ensuite la fermeture dorsale de l’embryon.
Le mésoderme se métamérise et se régionalise et donne alors, le mésoderme somatique (qui est à l’origine des muscles larvaires) et le mésoderme viscéral (qui est lui, à l’origine du tube digestif).
Les deux ébauches endodermiques de l’intestin moyen antérieur et postérieur se rejoignent vers la douzième heure. Les ébauches de la chaîne nerveuse ventrale vont se développer de plus en plus à l’avant avec les corps cérébroïdes. La chitine apparaît vers la treizième heure et les premiers mouvements musculaires vers la quatorzième.
E Les disques imaginaux.
L’ectoderme va former une sorte de sac aplati constitué d’une seule couche de cellules accrochées intérieurement à l’ectoderme de la larve sous l’influence de l’ecdysone (hormone de mue). Au moment de la métamorphose, ces disques imaginaux s’évaginent (se retournent comme des doigts de gants) pour prendre leur aspect apindiculaire définitif.
On distingue 19 disques dont :
- 3 paires pour la tête,
- 3 paires thoraciques-dorsales qui donnent les pro, méso, méta thorax avec les appendices correspondant (paire d’ailes et d’élytres),
- 3 paires thoraciques-ventrales qui donneront les trois paires de pattes,
- 1 disque génital impair.
Aucun commentaire jusqu'à présent.